En todos los seres vivos, cada gen interviene en la
realización de una función concreta. Muchos seres vivos tenemos dos dotaciones similares
de genes, procedentes cada una de los dos progenitores. Por tanto, tenemos
dos copias de cada gen. Por eso se dice que somos diploides, que quiere decir
“dos juegos”.
Mostrando entradas con la etiqueta Heterosis. Mostrar todas las entradas
Mostrando entradas con la etiqueta Heterosis. Mostrar todas las entradas
viernes, 31 de agosto de 2018
viernes, 23 de junio de 2017
¿Raza pura?
Según mi modo de ver, una raza es un conjunto de seres vivos que, dentro de una especie, poseen caracteres distintivos que se transmiten de padres a hijos. Esos caracteres pueden ser morfológicos, fisiológicos, de comportamiento, etc. En biología se discute sobre la naturaleza biológica de este concepto, y se define en términos ecológicos, como posible consecuencia de aislamiento geográfico.
En animales domésticos se tiende a asignar el calificativo de raza, como grupo, a un conjunto de individuos que, mediante selección artificial, ha ido acumulando caracteres que los distinguen del resto de individuos de su misma especie. Para lograrlo, no es raro recurrir a cruzamientos altamente consanguineos, de modo que la madre de un animal puede ser su hermana o su tía, por ejemplo. En los individuos pertenecientes a razas animales, aunque existe variabilidad, la morfología suele ser muy uniforme y debido a su ascendencia altamente consanguinea, suelen ser homocigotos para muchos de sus alelos. Esa alta proporción de homocigosis hace que sean individuos proclives a sufrir enfermedades u otros males causados por debilidad.
Fuera de este tema, y de modo general, debo indicar que la biología estudia fenómenos que ocurren en la naturaleza. Si por alguna razón, estamos en desacuerdo con lo que ocurre en el medio natural, debemos revisar nuestras ideas, pues la naturaleza no se equivoca. Los equivocados somos nosotros.
En una entrada anterior, comenté que los cruzamientos que se pueden producir entre individuos siempre generan una considerable proporción de homocigotos. Como este tipo de cruzamientos se repiten de modo constante a los largo de generaciones en poblaciones naturales, hemos de pensar que la proporción de homocigotos en la naturaleza aumenta de manera constante. Por eso se creía que en esas poblaciones el estado normal de los individuos era el de homocigosis, siendo el de heterocigosis un estado transitorio.
Por otra parte, existía un reto ante la comunidad científica y era el mayor rendimiento, en todos los sentidos, de los maíces híbridos. Se dijo que “en ellos” se daba una situación, aún por definir, a la que se llamó heterosis. No obstante, esa eficacia de los maíces híbridos decaía de modo fulminante luego de algunas generaciones de autofecundación.
A finales de la década de 1960, mediante técnicas nuevas de análisis, se encontró que en poblaciones naturales el número de heterocigotos es elevado y su proporción se mantiene a lo largo de las generaciones. Esto planteó dos tipos de preguntas, a las que se les buscó respuesta a lo largo de los años siguientes.
Las dos preguntas eran, por una parte, a qué se debía la presencia de tantos heterocigotos en esas poblaciones y, además, cómo se mantenía su frecuencia, siendo probado que al cruzarse generaban una elevada proporción de homocigotos.
Desde la década de 1940 ya se admitía que la acción primera de un alelo era la síntesis de un péptido . Cada gen regula una función debido al enzima cuya síntesis regula. Los diferentes alelos del gen regulan la misma función, pero con diferentes matices. Por ejemplo, supongamos que el alelo Aregula la síntesis del enzima A y el a, la enzima a. Siguiendo con el ejemplo, la enzima A permite vivir bien en unas condiciones determinadas de temperatura, y el a en otras ligeramente distintas. El homocigoto AA sólo tendrá enzima A y su rango ambiental estará restringido al determinado por ese enzima. Por su parte, el homocigoto aa tendrá un óptimo de temperatura ligeramente distinto, pues sólo tiene enzima a. Pero el heterocigoto Aa posee ambos enzimas y su rango vital, en cuanto a temperaturas se refiere, es más amplio que el de cualquier homocigoto AA o aa.
En geles resultantes de electroforesis se ven cómo los homocigotos presentan una sola banda, mientras que el heterocigoto presenta dos. Cada banda corresponde a un enzima. El antiguo y desconocido efecto de heterosis definía esta situación de mayor versatilidad vital de los heterocigotos. Cálculos teóricos realizados indican que todos los seres vivos somos heterocigotos para un importante número de genes.
Hay enzimas llamados monómeros porque están constituidos por una sola cadena. Los hay formados por dos cadenas, y los conocemos como dímeros. En los casos de enzimas dímeros, los homocigotos poseen un solo tipo de enzima AA o aa, pues los dos alelos que poseen son iguales y como consecuencia, las cadenas proteicas sintetizadas, también. En heterocigotos se presenta un hecho interesante, pues hay tres tipos de enzimas: AA, aa, y Aa, dándose la circunstancia de que uno de ellos, Aa sólo aparece en individuos heterocigotos.
 |
| Electroforesis de enzima dímero Los individuos 1 y 2 son homocigotos El individuo 3 es heterocigoto y presenta el enzima propio de su estado |
Tenemos aquí una explicación bioquímica del porqué la mayor frecuencia de heterocigotos en la naturaleza, su mayor riqueza enzimática y, por tanto, más versatilidad para adaptarse al medio.
La antigua heterosis que explicaba la mayor vitalidad de los maíces híbridos, la explicamos como consecuencia del alto grado de heterocigosis en esos maíces.
¿Qué decir, pues, de animales que constituyen razas puras? Tal vez sean el resultado de una intensa selección artificial, pero deben ser homocigotos para muchos de sus genes, lo cual explica su poca variabilidad. Pero esa poca variabilidad, junto al elevado grado de consanguinidad ejercida para obtener esa situación, puede hacer de ellos unos individuos muy poco resistentes ante cambios ambientales de cualquier tipo.
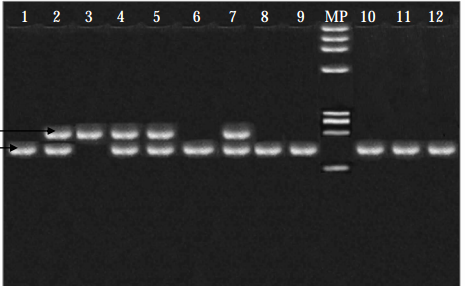 |
| Explicación en el texto |
En la figura presento el resultado de una elestrofóresis analizando de presencia de enzimas en varios individuos, numerados desde el 1 al 12 en la fila superior. Cada banda corresponde a un enzima y cada enzima está codificada por un alelo. Todos los casos han sido sometidos a las mismas condiciones físico-químicas.
Los individuos 1, 6, 8, 9, 10, 11 y 12 presentan un solo enzima que ha corrido poco. El individuo 3 presenta un solo enzima que, en igualdad de condiciones, ha corrido más. Los individuos 2, 4, 5 y 7 presentan dos enzimas: uno de ellos es como el que tiene el individuo 1, 6, 8, 9, 10, 11 y 12, el otro enzima es el propio del individuo 3.
Hay dos tipos de homocigotos, que reconocemos por tener un solo tipo de enzima, Los heterocigotos poseen los dos tipos.
Aquí los alelos se pueden denominar por su comportamiento en electroforesis, así, los alelos que están en 1, 6, y demás homozigotos reciben el nombre de "lentos", mientras que el que aparece en 3, es "rápido".
MP es un control de la marcha de los enzimas.
Los individuos 1, 6, 8, 9, 10, 11 y 12 presentan un solo enzima que ha corrido poco. El individuo 3 presenta un solo enzima que, en igualdad de condiciones, ha corrido más. Los individuos 2, 4, 5 y 7 presentan dos enzimas: uno de ellos es como el que tiene el individuo 1, 6, 8, 9, 10, 11 y 12, el otro enzima es el propio del individuo 3.
Hay dos tipos de homocigotos, que reconocemos por tener un solo tipo de enzima, Los heterocigotos poseen los dos tipos.
Aquí los alelos se pueden denominar por su comportamiento en electroforesis, así, los alelos que están en 1, 6, y demás homozigotos reciben el nombre de "lentos", mientras que el que aparece en 3, es "rápido".
MP es un control de la marcha de los enzimas.
viernes, 17 de febrero de 2017
Un encuentro inesperado
En todos los seres vivos, cada gen interviene en la realización de una función concreta. Muchos seres vivos tenemos dos dotaciones de genes, procedentes de cada uno de los dos progenitores. Por tanto, tenemos dos copias de cada gen. Por eso se dice que somos diploides, que quiere decir “dos juegos”.
De todos modos, las funciones se pueden realizar de modos diferentes. En nosotros mismos, conocemos múltiples tonalidades de color de ojos, de pelo, o diferentes grupos sanguíneos. En vegetales hay variabilidad en el color de las flores, en los bordes de las hojas o en el aspecto general de la planta, por citar tres casos. Los responsables de realizar las mismas funciones, pero de modos alternativos, reciben el nombre de “alelos”, que en griego significa “otro” y son variaciones de un mismo gen. Son alelos del mismo gen los que, por ejemplo, determinan nuestros diferentes grupos sanguíneos, los diversos colores de plantas cultivadas o los diferentes pelajes de algunos mamíferos.
Cuando un individuo tiene iguales los dos alelos de un gen, decimos de él que es “homocigoto”, si son diferentes, decimos que es “heterocigoto”. En griego, “homo” significa igual, “hetero”, diferente y “cigoto”, huevo. Los tiene iguales por haber heredado la misma copia de cada progenitor. Si son diferentes, es por haber recibido copias distintas.
Estos genes que se han recibido, se transmiten tal cual salvo mutación, (fenómeno infrecuente) a los descendientes mediante los gametos. Los homocigotos forman sus gametos iguales en relación al gen para el que lo son. Los heterocigotos forman dos tipos de gametos, con igual frecuencia y cada uno de ellos con uno de los dos alelos diferentes que posee.
Con esta introducción, que me está resultando larga, quiero sentar las bases de lo que voy a decir. Si representamos a un gen mediante una letra, por ejemplo, T y a su alelo con la misma letra, pero con grafía diferente, t, tendremos dos tipos de homocigotos, TT y tt y un solo tipo de heterocigoto, Tt.
Los primeros estudios rigurosos sobre herencia los realizó y publicó Mendel hace más de un siglo y medio (1866), aunque no fue comprendido más que a partir de 1900. Curiosamente, todo tipo de posible cruzamiento que se realiza con heterocigotos, genera una descendencia con abundancia de homocigotos, hasta un 50% del total de la progenie. Lógicamente, cuando se cruzan homocigotos similares, todos los descendientes son también homocigotos. Esto llevó a pensar a los científicos de entonces que en la Naturaleza, donde los cruzamientos se realizan al azar, debería haber una gran cantidad de homocigotos y pocos heterocigotos. Éstos, los heterocigotos, vendrían a ser como situaciones genéticas inestables, en cuya descendencia aparecían abundantes homocigotos, siendo por tanto difícil de mantener presentes en una población a lo largo de las generaciones. Este planteamiento generó un fecundo cuerpo de conocimientos, que hoy conocemos bajo el nombre de “genética clásica”.
 |
| EL GRAN LOGRO DE LA GENETICA CLÁSICA |
Lógicamente, puesto que los homocigotos eran los más frecuentes en las poblaciones naturales, tendrían que ser los más adaptados a sus ambientes. Había una excepción inexplicable: el caso de los maíces híbridos. Se les atribuía “superdominancia” sin saber lo que eso significaba, o bien “heterosis”, algo relacionado con la mejor situación de los heterocigotos, concepto que en parte se mantiene en la actualidad.
Cuando nuestras predicciones teóricas están en discordancia con lo que encontramos en la Naturaleza, debemos revisar nuestras predicciones con el convencimiento de que la Naturaleza no se equivoca. Y más convencidos aún si realizamos constataciones en diversas situaciones con similares resultados.
He dicho en otras ocasiones que nuestros conocimientos están en constante revisión. Esa revisión sirve para mantenerlos o rechazarlos si acaso no superan las pruebas que se puedan hacer aplicando nuevas técnicas que se vayan descubriendo.
Eso es lo que, en 1966, hicieron dos investigadores. R. Levontin y J.L. Hubby publicaron unos resultados inesperados, pues rompía todo cuanto se creía según la Genética clásica. Aplicando métodos de electroforesis en seres procedentes de poblaciones naturales, encontraron más heterocigotos de los que cabría esperar según los razonamientos teóricos. Las pruebas se repitieron estudiando otras especies, y los resultados fueron coincidentes. Más heterocigotos de lo esperado.
Esto presentó ante los investigadores un doble aspecto. Unos, los poco emprendedores, se dejaron vencer por la evidencia sin mayores alicientes, y abandonaron sus estudios. Para la mayoría, no obstante, estos resultados indicaban la falsedad de la teoría anterior, que llevaba a la idea de la alta presencia de homocigotos en las poblaciones naturales. Este hecho se transformó en un reto para la comunidad científica y era precisa la búsqueda de respuestas adecuadas. Se planteaban dos preguntas importantes: 1º, por qué habían más heterocigotos de lo esperado y 2º, cómo se mantenían en altas frecuencias si en sus descendencias aparecían elevadas proporciones de homocigotos.
Fue una época apasionante aquella de buscar esas respuestas, y otras que fueron surgiendo. También aparecieron nuevos conceptos de selección natural, y se observaron diversos tipos suyos: selección estacional, selección dependiente de frecuencias, selección disruptiva, etc. etc.
Seguimos en eso, pero he querido relatar, mediante este ejemplo, vivido por mí y recordado con cariño por corresponder a una época fecunda de mi trabajo, cómo la ciencia avanza desechando conceptos obsoletos y buscando respuestas a las constantes preguntas tipo ¿cómo? ¿por qué? Y similares.
En muchas ocasiones los mismos errores suelen ser muy fecundos en información científica. Su última contribución a la ciencia es mostrar su falsedad. Y, recuerdo, las preguntas han de estar bien formuladas y en sus momentos adecuados.
Suscribirse a:
Entradas (Atom)