Algunos amigos me comentan que, al hablar de poblaciones
pequeñas, me acerco más a ideas de M. Kimura que a las de Darwin. No les falta
razón, pero han tenido que hacerme reflexionar sobre eso. Llevo tantos años
dándole la vuelta a las mismas cosas, que ya no soy capaz de reconocer su
paternidad.
Mostrando entradas con la etiqueta Electroforesis. Mostrar todas las entradas
Mostrando entradas con la etiqueta Electroforesis. Mostrar todas las entradas
viernes, 13 de septiembre de 2019
viernes, 23 de junio de 2017
¿Raza pura?
Según mi modo de ver, una raza es un conjunto de seres vivos que, dentro de una especie, poseen caracteres distintivos que se transmiten de padres a hijos. Esos caracteres pueden ser morfológicos, fisiológicos, de comportamiento, etc. En biología se discute sobre la naturaleza biológica de este concepto, y se define en términos ecológicos, como posible consecuencia de aislamiento geográfico.
En animales domésticos se tiende a asignar el calificativo de raza, como grupo, a un conjunto de individuos que, mediante selección artificial, ha ido acumulando caracteres que los distinguen del resto de individuos de su misma especie. Para lograrlo, no es raro recurrir a cruzamientos altamente consanguineos, de modo que la madre de un animal puede ser su hermana o su tía, por ejemplo. En los individuos pertenecientes a razas animales, aunque existe variabilidad, la morfología suele ser muy uniforme y debido a su ascendencia altamente consanguinea, suelen ser homocigotos para muchos de sus alelos. Esa alta proporción de homocigosis hace que sean individuos proclives a sufrir enfermedades u otros males causados por debilidad.
Fuera de este tema, y de modo general, debo indicar que la biología estudia fenómenos que ocurren en la naturaleza. Si por alguna razón, estamos en desacuerdo con lo que ocurre en el medio natural, debemos revisar nuestras ideas, pues la naturaleza no se equivoca. Los equivocados somos nosotros.
En una entrada anterior, comenté que los cruzamientos que se pueden producir entre individuos siempre generan una considerable proporción de homocigotos. Como este tipo de cruzamientos se repiten de modo constante a los largo de generaciones en poblaciones naturales, hemos de pensar que la proporción de homocigotos en la naturaleza aumenta de manera constante. Por eso se creía que en esas poblaciones el estado normal de los individuos era el de homocigosis, siendo el de heterocigosis un estado transitorio.
Por otra parte, existía un reto ante la comunidad científica y era el mayor rendimiento, en todos los sentidos, de los maíces híbridos. Se dijo que “en ellos” se daba una situación, aún por definir, a la que se llamó heterosis. No obstante, esa eficacia de los maíces híbridos decaía de modo fulminante luego de algunas generaciones de autofecundación.
A finales de la década de 1960, mediante técnicas nuevas de análisis, se encontró que en poblaciones naturales el número de heterocigotos es elevado y su proporción se mantiene a lo largo de las generaciones. Esto planteó dos tipos de preguntas, a las que se les buscó respuesta a lo largo de los años siguientes.
Las dos preguntas eran, por una parte, a qué se debía la presencia de tantos heterocigotos en esas poblaciones y, además, cómo se mantenía su frecuencia, siendo probado que al cruzarse generaban una elevada proporción de homocigotos.
Desde la década de 1940 ya se admitía que la acción primera de un alelo era la síntesis de un péptido . Cada gen regula una función debido al enzima cuya síntesis regula. Los diferentes alelos del gen regulan la misma función, pero con diferentes matices. Por ejemplo, supongamos que el alelo Aregula la síntesis del enzima A y el a, la enzima a. Siguiendo con el ejemplo, la enzima A permite vivir bien en unas condiciones determinadas de temperatura, y el a en otras ligeramente distintas. El homocigoto AA sólo tendrá enzima A y su rango ambiental estará restringido al determinado por ese enzima. Por su parte, el homocigoto aa tendrá un óptimo de temperatura ligeramente distinto, pues sólo tiene enzima a. Pero el heterocigoto Aa posee ambos enzimas y su rango vital, en cuanto a temperaturas se refiere, es más amplio que el de cualquier homocigoto AA o aa.
En geles resultantes de electroforesis se ven cómo los homocigotos presentan una sola banda, mientras que el heterocigoto presenta dos. Cada banda corresponde a un enzima. El antiguo y desconocido efecto de heterosis definía esta situación de mayor versatilidad vital de los heterocigotos. Cálculos teóricos realizados indican que todos los seres vivos somos heterocigotos para un importante número de genes.
Hay enzimas llamados monómeros porque están constituidos por una sola cadena. Los hay formados por dos cadenas, y los conocemos como dímeros. En los casos de enzimas dímeros, los homocigotos poseen un solo tipo de enzima AA o aa, pues los dos alelos que poseen son iguales y como consecuencia, las cadenas proteicas sintetizadas, también. En heterocigotos se presenta un hecho interesante, pues hay tres tipos de enzimas: AA, aa, y Aa, dándose la circunstancia de que uno de ellos, Aa sólo aparece en individuos heterocigotos.
 |
| Electroforesis de enzima dímero Los individuos 1 y 2 son homocigotos El individuo 3 es heterocigoto y presenta el enzima propio de su estado |
Tenemos aquí una explicación bioquímica del porqué la mayor frecuencia de heterocigotos en la naturaleza, su mayor riqueza enzimática y, por tanto, más versatilidad para adaptarse al medio.
La antigua heterosis que explicaba la mayor vitalidad de los maíces híbridos, la explicamos como consecuencia del alto grado de heterocigosis en esos maíces.
¿Qué decir, pues, de animales que constituyen razas puras? Tal vez sean el resultado de una intensa selección artificial, pero deben ser homocigotos para muchos de sus genes, lo cual explica su poca variabilidad. Pero esa poca variabilidad, junto al elevado grado de consanguinidad ejercida para obtener esa situación, puede hacer de ellos unos individuos muy poco resistentes ante cambios ambientales de cualquier tipo.
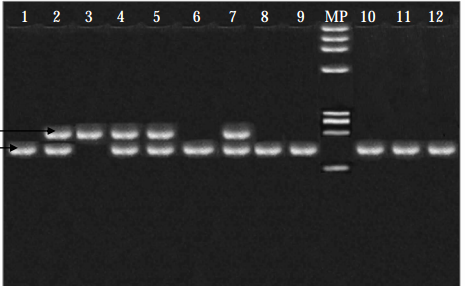 |
| Explicación en el texto |
En la figura presento el resultado de una elestrofóresis analizando de presencia de enzimas en varios individuos, numerados desde el 1 al 12 en la fila superior. Cada banda corresponde a un enzima y cada enzima está codificada por un alelo. Todos los casos han sido sometidos a las mismas condiciones físico-químicas.
Los individuos 1, 6, 8, 9, 10, 11 y 12 presentan un solo enzima que ha corrido poco. El individuo 3 presenta un solo enzima que, en igualdad de condiciones, ha corrido más. Los individuos 2, 4, 5 y 7 presentan dos enzimas: uno de ellos es como el que tiene el individuo 1, 6, 8, 9, 10, 11 y 12, el otro enzima es el propio del individuo 3.
Hay dos tipos de homocigotos, que reconocemos por tener un solo tipo de enzima, Los heterocigotos poseen los dos tipos.
Aquí los alelos se pueden denominar por su comportamiento en electroforesis, así, los alelos que están en 1, 6, y demás homozigotos reciben el nombre de "lentos", mientras que el que aparece en 3, es "rápido".
MP es un control de la marcha de los enzimas.
Los individuos 1, 6, 8, 9, 10, 11 y 12 presentan un solo enzima que ha corrido poco. El individuo 3 presenta un solo enzima que, en igualdad de condiciones, ha corrido más. Los individuos 2, 4, 5 y 7 presentan dos enzimas: uno de ellos es como el que tiene el individuo 1, 6, 8, 9, 10, 11 y 12, el otro enzima es el propio del individuo 3.
Hay dos tipos de homocigotos, que reconocemos por tener un solo tipo de enzima, Los heterocigotos poseen los dos tipos.
Aquí los alelos se pueden denominar por su comportamiento en electroforesis, así, los alelos que están en 1, 6, y demás homozigotos reciben el nombre de "lentos", mientras que el que aparece en 3, es "rápido".
MP es un control de la marcha de los enzimas.
martes, 12 de abril de 2016
Reflexiones sobre el azar
+Laura Villoria me comenta que, al hablar de poblaciones pequeñas, me acerco más a ideas de M. Kimura que a las de Darwin. No le falta razón, pero ha tenido que hacerme reflexionar sobre eso. Llevo tantos años dándole la vuelta a las mismas cosas, que ya no soy capaz de reconocer su paternidad.
Kimura no es muy conocido para el gran público. Sólo Darwin, por el brutal comentario, falsamente atribuido a él, acerca de la supervivencia del más fuerte, permanece en el sentir popular propiciado por gente matona, inculta y satisfecha de sí. (Me gustaría saber cuántos españoles conocen el trabajo de Cajal, el que le hizo merecer el Premio Nobel).
 |
| VARIABILIDAD GENÉTICA ENZIMATICA. CADA VERTICAL REPRESENTA UN INDIVIDUO MUESTREADO |
En la década de 1960, en genética, las ideas parecían estar consolidadas. Fue entonces cuando apareció una nueva técnica de análisis enzimático, electroforesis en gel, y dos investigadores, Hubby y Lewontin, la aplicaron a muestras de individuos procedentes de muy diversas poblaciones naturales. Los resultados fueron espectaculares por lo inesperados. Había mucha variabilidad genética no visible en las poblaciones, mucha más de la explicable con los criterios de entonces.
Lo he dicho en otras entradas de este blog. La ciencia busca explicar el entorno con las herramientas de que dispone. Con estas explicaciones se construye un cuerpo doctrinal, nunca cerrado, con el que se pretende interpretar y responder a las preguntas que se vayan formulando. Para comprobar la veracidad de las hipótesis utilizadas, se vuelven a comprobar cuando se dispone de nuevos métodos de análisis.
Para comprobar las ideas genéticas imperantes, en 1966, se aplicaron las técnicas de la electroforesis a las poblaciones naturales. Las ideas anteriores sufrieron un tremendo revulsivo, a la vez que aparecieron nuevas preguntas por contestar, muchas de ellas aún en vías de resolución.
 |
| VEMOS UN GRAN VARIABILIDAD ENTRE LOS INDIVIDUOS MUESTREADOS |
Como apareció mucha variabilidad génica, es decir más de la esperada, las preguntas giran alrededor de este dato. ¿Cómo surge esta variabilidad? Y más importante aún, ¿cómo se mantiene? Cómo aparece la variabilidad está claro que es por mutación, pero lo esperado sería que se perdiese al poco de aparecer. No obstante, hay mucha en las poblaciones naturales. ¿Cómo se mantiene? Es decir, cómo es que la selección natural no va eliminando, a la velocidad que sea, toda la variabilidad que no genere mayor adaptación a sus portadores.
Porque, y hay que tenerlo en cuenta, mucha variabilidad requiere poblaciones grandes, pero en la naturaleza las poblaciones no lo son. Más bien, tienen tamaños limitados. En una bandada de aves, en un pequeño bosque o en una colonia de celentéreos, es donde se llevan a cabo los procesos evolutivos que he comentado en mas de una ocasión. Y no tienen grandes tamaños. Para comprender lo que ocurría, se estudiaron diversos procesos biológicos y aparecieron casos de selección explicables mediante las teorías de Darwin, pero aplicados a casos concretos, como la selección estacional, la dependiente de frecuencias, la gamética, etc. etc.
Aquí había un fallo conceptual, grande. Nadie quería abandonar la idea de la selección darwiniana, el pensar que cada gen, a su modo, contribuye al valor selectivo de su portador. ¿Sumando o multiplicando? Es imposible que todos los genes actúen por igual, pues los letales, por ejemplo, tendrían un efecto enmascarador de los demás, matando a su portador.

Es en esta situación de la contribución de los diferentes genes al valor de un individuo frente a la selección, cuando Kimura propone que, al igual que la contribución de los genes a ese valor es variable, tambien los puede haber con valor 0, es decir, que sean neutros ante la selección. Así, unos serán beneficiados, otros rechazados y otros, la mayoría, serán neutros ante la selección. Esto explicaría la gran cantidad de variabilidad encontrada, y cuya explicación resultaba muy difícil desde un punto de vista adaptativo. Simplemente, porque al ser neutros, la selección no los “ve”.
Me gustan mucho estas ideas, pues a veces vamos más allá de lo que nos dicen nuestros estudios. Por ejemplo, a veces queremos saber qué ve la selección natural, y no nos damos cuenta de que es una tendencia sin mayor trascendencia a corto plazo. Que depende mucho de las condiciones ambientales y que de ese modo, sus individuos seleccionados pueden ser muy diferentes a lo largo de las generaciones. A veces he oído decir “lo que tendría que ocurrir…” y me he reído, porque parece que haya quienes quieran darle la vuelta a las cosas. Nosotros estudiamos la naturaleza, no tenemos que indicarle lo que debe hacer.
Una cosas derivada de todo esto es que (pensábamos) los gametos formadores de una generación, lo eran gracias a haber sido favorecidos por diversos aspectos de la selección. ¿Y si eran neutros, si la selección no los veía? Entonces entraba en escena el azar.
No todos los genes son selectivos ni todos son neutros. Desconozco cómo se integra el valor de cada gen en el valor general del gameto, pero algo de esto hay.
Antes de Kimura, las poblaciones genéticas se definían como “conjunto infinito de individuos…” Luego, con los pies más en el suelo, tal vez comprendiendo que no existe una población de tamaño infinito, se define como “conjunto de individuos que se cruzan entre si…” Ese es el objeto de la evolución, la población que de mayor o menor tamaño, siempre en pequeña en términos genéticos, y sujeta a diversos efectos selectivos, Pero también al azar.

Un error conceptual evidente (las poblaciones son infinitas…) que no supimos ver hasta pasado mucho tiempo. Con frecuencia me pregunto con cuántos errores conceptuales de este tipo estaremos trabajando. El aceptar la idea de Kimura, suponía para muchos una especie de rechazo a Darwin, por eso tardó tanto en serlo. En aquel tiempo yo hacía mi tesis doctoral. Desde entonces, he visto que a las personas mayores les cuesta adoptar ideas nuevas, tal vez pensando traicionar a antiguos maestros. No lo sé. Creo que no participo de ese modo de pensar.
Gracias +Laura Villoria, por hacerme reflexionar sobre esto.
Suscribirse a:
Entradas (Atom)